

Descargar este artículo en formato PDF
Bozarth, Steven y Richard D. Hansen
2001 Estudios paleobotánicos de Nakbe: Evidencias preliminares de ambientes y cultivos en el Preclásico. En XIV Simposio de Investigaciones Arqueológicas en Guatemala, 2000 (editado por J.P. Laporte, A.C. Suasnávar y B. Arroyo), pp.369-382. Museo Nacional de Arqueología y Etnología, Guatemala (versión digital).
26
ESTUDIOS PALEOBOTÁNICOS DE NAKBE:
EVIDENCIAS PRELIMINARES DE AMBIENTE Y CULTIVOS EN EL PRECLÁSICO
Steven Bozarth
Richard D. Hansen
Nota de la edición: plática presentada en el XII Simposio de Investigaciones Arqueológicas en Guatemala, 1998
La identificación de los antiguos restos botánicos dentro de un sitio arqueológico se ha convertido en una importante contribución al interpretar la dinámica y el proceso cultural de sociedades complejas tempranas. Estos estudios se han logrado llevar a cabo gracias al creciente interés no solo en recobrar el polen antiguo, los isótopos y otros restos vegetales, sino en realizar estudios de botánica que ayuden a establecer información sobre paleo-ambiente, los cultivos y la ecología antigua.
Se han llevado a cabo observaciones preliminares sobre el polen, los isótopos, los tipos y la naturaleza de las comunidades vegetales en Nakbe por parte de la Universidad de Texas A&M, la Universidad de San Carlos de Guatemala, la Universidad de California, Berkeley, y la Universidad de California, Los Angeles (Jones 1991; Weinstein 1994, Castañeda y Castañeda 1994; Hansen 1995; Romero 1996; Castañeda 1997; Romero y Schreiner 1998), anticipándose a investigaciones mucho más complejas que se puedan dar en un futuro con acercamientos multidisciplinarios para estudiar el medio ambiente de la antigüedad, el clima, la dieta, y la respuesta humana a los retos y las oportunidades dadas por el medio ambiente. Es notable el potencial de tales estudios para ayudar a establecer unas vistas diacrónicas de las actividades tantas climatológicas como culturales (Deevey et al. 1979; Brenner et al. 1990). Por ejemplo, en su importante estudio de 1979, Deevey et al., indicó la relación del polen con la afluencia de agua a los lagos de Yaxha y Sacnab (Figura 1).
Una de las observaciones notables es la reducción de polen arbóreo para finales del Preclásico Tardío, principios del Clásico Temprano y finales del Clásico Tardío. Esta reducción también corresponde al aumento de sedimentos en los lagos. Es posible, en base de estas correspondencias, de establecer la hipótesis que la deforestación pueda estar relacionada con la mayor concentración humana en estas épocas, y posiblemente hubo consecuencias ambientales por causa de tal interacción (Hansen 1994, 1995).
En la zona de la Cuenca Mirador, al norte de Petén, hay indicaciones de una extraordinaria estratigrafía en los bajos que se encuentran alrededor de los grandes sitios. Las investigaciones de John Jacob (1994) en las cercanías de Nakbe, revelaron una capa de paleo-suelos (Figura 2) enterrada bajo una capa de sedimentos de arcilla Maya. Un análisis de los isótopos sugirió que la capa del paleo-suelo consistió de plantas C-4, como maíz y zacate, mientras la capa de sedimento consistió de plantas C-3, o sea la vegetación del bosque actual (Figura 3).
Hansen ha interpretado esta capa como evidencia de un cambio ecológico debido a la sedimentación profunda, los cambios en los isótopos, la preservación del polen del zacate y de las hierbas que se dan en zonas húmedas en los sibales actuales, y la presencia de terrazas artificiales construidas del barro semejante al paleo-suelo enterrado en los bajos (Hansen 1994, 1995). Domínguez y Folan señalaron la misma intensidad de sedimentación en el área residencial de Calakmul, fechado para el Clásico Temprano, marcada por esta gran capa de arcilla (Domínguez 1993). De la misma forma, la evidencia de los grandes sibales indican los posibles restos fósiles del medio ambiente original de los bajos (Hansen 1994, 1995). Una muestra preliminar fue obtenida a una profundidad de 5.60 m por Jacob en la aguada Zacatal (Hansen 1994). Los análisis de polen hechos por el Dr. Eri Weinstein de la Universidad de Texas A&M, y la Dra.
Barbara Leyden de la Universidad del Sur de Florida, revelaron la rica preservación de toda la columna vertical del polen de zacate, plantas de medio húmedo y otras especies provenientes de pantanos húmedos (Weinstein 1993; Leyden 1994; Hansen 1995). Debido al éxito de la muestra obtenida por Jacob, Leyden y Weinstein, el Dr. Roger Byrne y Tom Schreiner de la Universidad de California en Berkeley, iniciaron investigaciones en la aguada Zacatal durante la temporada de 1998. Las investigaciones de Schreiner revelaron una modificación artificial de la orilla y piso de la aguada, realizada por los Mayas.
El fechamiento de esta modificación ya está en proceso de ser analizado por la Universidad de California en Berkeley. Las muestras cilíndricas obtenidas por Schreiner nos darían la posibilidad de analizar la historia ambiental de la zona. La evolución de los pantanos húmedos hacia un bajo en la actualidad es uno de los fenómenos de importancia porque se puede observar la transformación actual en el norte de Petén.
En ambientes tropicales es difícil la recuperación de polen porque los periodos de humedad y sequía ocasionan su destrucción. Sin embargo, en vista de estas dificultades, hemos recuperado muestras preservadas de contextos fechados para el Preclásico Medio (ca. 600 AC – 1000 AC) en Nakbe. Un entierro de la plataforma de la Estructura 32, fechando al Preclásico Medio, fue excavado por Francisco López, y reveló nueve clases de entidades taxonómicas, incluyendo el maíz (Zea mays). Un depósito excavado por Susan Snow (UCLA) en la Estructura 53, que también fecha para el Preclásico Medio, reveló una gran variedad de especies agrícolas incluyendo maíz, calabaza, chicozapote, palmera (escoba), pino, copo, chicem, ceiba, chacaj (Burcera simaruba), zacates, y posiblemente una muestra de cacao (Jones 1991; Hansen 1995:72).
Debido a los problemas que surgen al momento de recuperar una muestra adecuada de polen en las tierras bajas tropicales, el proyecto PRIANPEG deseaba saber si las fitolitas de ópalo en Nakbe se encontraban bien preservadas y en concentraciones adecuadas para ser analizados. Estudios previos efectuados por John Jones (1991) sugirieron la posibilidad de preservación de polen y de fitolitas. El hallazgo de un sistema de terrazas y modificaciones artificiales en Nakbe dio la oportunidad de iniciar tal estudio en un contexto de los sistemas agrícolas. Un objetivo fue dar acceso a la identificación de cultivos y plantas silvestres comestibles que pudieran estar presentes.
FORMACIONES FITOLÍTICAS
Las plantas tropicales que están en crecimiento absorben típicamente el agua que contiene sílice disuelto. Los cuerpos microscópicos de sílice se forman entonces por la silicificación parcial o total de las células de las plantas, las paredes celulares y los espacios intercelulares. Estos cuerpos de sílice resultantes, con formas y tamaños característicos son llamados fitolitas de ópalo (Wilding y Dress 1971). El término fitolita se deriva del griego phyton que significa planta, y lithos, que significa piedra. Ópalo es el nombre común del dióxido de sílice amorfo e hidratado.
La mayoría de las plantas están formadas por fitolitas que se producen en una gran variedad de formas y tamaños. Pueden ser característicos cuando su forma y/o tamaño es específico para un tipo particular de planta. Afortunadamente, muchas fitolitas son resistentes al clima y se preservan en la mayoría de suelos y sedimentos durante largos periodos de tiempo.
ESTABILIDAD DE LAS FITOLITAS
El proceso de disolución y estabilidad de fitolitas en los sedimentos no ha sido totalmente entendido. Los experimentos de laboratorio demuestran, sin embargo, que la solubilidad de las fitolitas se da en función de la forma y el tamaño de la partícula, su pH y la presencia de iones metálicos y/o sesquióxidos.
El deterioro químico ocurre en las superficies de las fitolitas. Los que son de paredes delgadas, como aquellos producidos en las hojas caídas de los árboles son altamente frágiles y susceptibles a deteriorarse debido a su configuración geométrica que da lugar a una gran área de superficie (Wilding y Drees 1971, 1974). La disolución de ópalo aumenta cuando la dimensión de la partícula decrece (Wilding et al. 1977, 1979), lo cual indica que las fitolitas pequeñas no se preservan en los sedimentos tanto como las fitolitas de mayor tamaño.
La mayoría de los reportes demuestran que la solubilidad del sílice es independiente del pH dentro de un rango de valores de 3 hasta debajo de 9, pero aumenta rápidamente con un pH superior a estos valores (Wilding et al. 1977). La estabilidad del ópalo también está dada en función de la presencia de ciertos iones metálicos (e.g, Al, Fe, Mg, Ca, etc), y sesquióxidos (óxidos en los que tres átomos de oxígeno o sus equivalentes se combinan con dos de cualquier otro elemento o radical). La absorción de iones de Al y Fe sobre la superficie del ópalo hará decrecer el índice de disolución del sílice debido a la formación de capas silíceas relativamente insolubles.
INVESTIGACIÓN DE FITOLITAS ARQUEOLÓGICAS
Una considerable cantidad de estudios sobre fitolitas se ha llevado a cabo en los neotrópicos. La mayor parte de estos trabajos se ha realizado sobre la agricultura precolombina, no obstante que algunos paleo-ambientes han sido reconstruidos en muchas áreas. La investigación de fitolitas relacionadas con la agricultura precolombina y la subsistencia de las plantas están todavía en su infancia en las Tierras Bajas Mayas, pero las investigaciones en Nakbe nos dieron la oportunidad de iniciar estudios correspondientes.
Muchas de las investigaciones mencionadas se han enfocado en el maíz, su importancia en muchas culturas precolombinas y la abundancia de fitolitas distintivos producidos en estas especies. Los de forma cruzada, característicos del maíz, han sido estudiados extensamente ya que son producidos comúnmente por esta planta. Aunque es menos común, también se forman en la subfamilia panicular de las hierbas y en algunas especies de la subfamilia del bambú. Este tipo de fitolita consiste en tres o cuatro lóbulos adheridos a un cuerpo central fácil de identificar (Piperno 1988).
El primer estudio arqueológico sobre las fitolitas de maíz fue realizado por Pearsall (1978). Las fitolitas de las hojas de muchas especies del maíz (hierbas paniculares), fueron comparadas con las de las hojas de tres hierbas paniculares silvestres nativas del Ecuador. Solo el maíz produjo fitolitas cruzadas grandes y extra grandes. Las medidas se basaron en dimensiones de eje corto (i.e., ancho). Las cruzadas se definieron como aquellos que miden 9 µm de largo más que de ancho. Esta diferencia en el tamaño permitió una identificación tentativa de las fitolitas del maíz aisladas del sedimento fechado para 2450-2150 AC y 1500 AC en sitios de Ecuador (Pearsall 1978).
Investigaciones subsecuentes realizadas por Pearsall (1982) sobre una colección mucho más extensa de hierbas paniculares (62 especies) nativas del suroeste del Ecuador, demostraron que cinco entidades taxonómicas producen formas cruzadas de mayor tamaño. De cualquier forma, estas entidades produjeron únicamente una frecuencia baja de formas cruzadas grandes. El ensamblaje total de fibras cruzadas en las especies de las hierbas fue distinto a la producida en el maíz. La presencia de fósiles de fitolitas de forma cruzada apoya la hipótesis de que el maíz estaba presente en el sitio Real Alto, tal como lo reportó Pearsall en 1978.
Los análisis de las fitolitas del maíz fueron perfeccionados por Piperno (1984), quien demostró que existen tres criterios para la identificación de las mismas. Estos son: (1) altos porcentajes de fitolitas cruzadas de gran tamaño, (2) morfología tridimensional, y (3) rangos bajos que varían desde la forma de la pesa hasta los de forma cruzada. Las fitolitas que satisfacen el criterio 1 y 2 pueden identificarse como maíz.
Basándose en estos criterios, el maíz fue identificado en dos sitios precolombinos de Panamá: Cueva de los Ladrones (4919-1820 AC), y en el refugio de Aguadulce (ca. 2100-1000 AC). La presencia de polen de maíz en Ladrones aportó evidencias para apoyar los datos de fitolitas (Piperno 1984; Piperno et al. 1985). También se recuperaron fitolitas cruzadas de maíz en el Lago Ayauch, al sureste de la Amazonia Ecuatoriana, en sedimentos que posiblemente se fecharon para 6000 AC (Bush et al. 1989; Piperno 1990).
Las mazorcas son parte particularmente importante de la planta del maíz al estudiar las características, ya que éstas, con o sin cáscara, eran sin duda llevadas a las aldeas precolombinas para desgranar. Posterior a la remoción de los granos, las mazorcas serían desechadas dentro o alrededor del sitio, o pudieron haber sido usadas como fuentes de combustible. Por lo tanto, es fácil encontrar las de las mazorcas en los basureros de los sitios precolombinos en donde se cultivaba el maíz.
No se conocen estudios fitolíticos sobre mazorcas de maíz de variedades provenientes de los cultivos de Centro y Sur América. De cualquier forma, la investigación de fitolitas en variedades tradicionales, no híbridas de maíz provenientes de las Grandes Planicies, y las especies de referencia nativas a la misma región, determinaron que las mazorcas producen numerosas fitolitas taxonómicamente valiosas (Bozarth 1987a, 1989a, 1989b). Investigaciones posteriores concluyeron que las mazorcas producen un tipo único de fitolita. Este tipo, común dentro de las cuatro variedades estudiadas, tiene las siguientes características: la parte superior (que es más estrecha que la parte inferior), y la parte inferior no son sinuosas; la parte inferior tiene bordes redondeados (vistos desde arriba o abajo): y la parte inferior es plana, la parte superior es plana u ondulada, los lados son distintivamente cóncavos, y la parte superior e inferior tienen la misma dimensión o la primera es más larga que la última (cuando se observa de lado). La evidencia macrofósil apoya esta conclusión (Bozarth 1989b, 1993a).
Investigaciones taxonómicas sistemáticas similares realizadas por Bozarth (1992a, 1992b 1994, 1996, 1997) sobre el crecimiento de plantas cultivadas en el Suroeste americano y la flora nativa a la misma región, concluyeron que las mazorcas, de las cinco clases de maíz, producen fitolitas similares, y aparentemente características de esta planta. Las fitolitas de las mazorcas se preservan bien generalmente y se han recuperado en 24 sitios prehistóricos de Arizona. Tomados en conjunto, los estudios sistemáticos de las fitolitas de las mazorcas indican que los mismos tipos de fitolitas son probablemente propias del maíz en los sitos precolombinos de Mesoamérica.
También se forman en el maíz otras fitolitas taxonómicamente valiosas. Mulholland (1987) reportó que los que tienen forma de pesa con acanaladuras formadas en las hojas y en las cáscaras de dos variedades de maíz de Dakota del Norte eran indicadores potenciales del maíz en sitios arqueológicos. Un tipo acanalado particular, común en la porción aérea del maíz, con extremos redondeados y/o puntiagudos fue reportado por Bozarth (1992 a, 1992b, 1994, 1996, 1997), como es propio del maíz en Arizona. Al igual que las fitolitas de las mazorcas, los bilobulados acanalados se preservan bien en la mayoría de sedimentos y han sido identificados en varios sitios de Arizona.
El primer estudio fitolítico de la calabaza se realizó sobre la corteza de especies seleccionadas de C.maxima y C.pepo, así como C.foetidissima (calabaza silvestre). Los análisis de las fitolitas de cucúrbita demostraron que las fitolitas esféricas y hemisféricas son superficies profundamente onduladas y con cavidades continuas, se producen con una frecuencia mayor en la calabaza que en el calabacín búfalo silvestre y pueden ser utilizados como indicadores de la calabaza en los Estados Unidos centrales (Bozarth 1985, 1986, 1987b).
Estudios posteriores demostraron que otra especie de calabaza, la C.mixta, también producía fitolitas esféricas y onduladas en su corteza. Adicionalmente, los cuerpos recto-curvilíneos elongados con secciones circulares u ovales cruzados y superficies onduladas se forman en las cortezas de variedades selectas de C.mixta, C.moshata y C.pepo. Otro tipo, con cinco a siete gruesas traqueidas, se forma comúnmente en la pulpa de C.maxima, C.mixta y C.moshata. Los primeros dos tipos de fitolitas parecen ser características de la calabaza del centro de Arizona y las Grandes Planicies. Las traqueidas de cinco a siete lados se encontraron solamente en cucúrbita y lagenaria (Bozarth 1992b, 1994). Sumado a esto, las fitolitas taxonómicamente valiosas se forman en las hojas de calabaza. De cualquier modo, ninguna se encuentra sólidamente silicificada, indicando que probablemente no se preservan en el sedimento (Piperno 1988).
Existe un considerable potencial para la documentación arqueológica de la calabaza usando las fitolitas de la corteza ya que las frutas se habrían llevado a las aldeas precolombinas (Piperno 1985). Estas fitolitas se han recuperado dentro de muestras sedimentarias recolectadas en dos aldeas prehistóricas de las Grandes Planicies centrales (Bozarth 1985, 1986, 1987b), y en el suroeste americano (Bozarth 1997). También se han recuperado fitolitas de calabaza en sedimentos arqueológicos de Panamá de hasta 3000 años de antigüedad (Piperno 1985).
Calabazas de botella y cucharón (lagenaria siceraria) son también parte de la familia de la calabaza (cucurbitaceae). El análisis inicial de estas calabazas indicó que las cortezas de estas plantas domesticadas producían únicamente fitolitas onduladas superficiales (Bozarth 1985, 1987b). De cualquier forma, los análisis posteriores en otras calabazas domesticadas demostraron que también se manifiestan las fitolitas grandes, onduladas y globulares. Estos difieren de las fitolitas propias de la calabaza en que sus áreas onduladas son mayores, las superficies pueden ser granuladas, y no son esféricas. En las cortezas también aparecen placas planas granuladas con bordes angulares curvilíneos. Las gruesas traqueidas espirales de cinco lados aparecen en la pulpa. Los dos primeros tipos de fitolitas parecen ser exclusivas de las especies domesticadas del centro de Arizona. Las traqueidas de cinco lados se observan únicamente en la cucúrbita y lagenaria (Bozarth 1994, 1996). Células fitolíticas distintivas con hebras segmentadas se forman en las hojas de la lagenaria siceraria (calabaza de botella). No obstante, fitolitas similares se observan en otros géneros de Cucurbitaceae (Piperno 1998).
Los frijoles cultivados (Phaseolus spp.) fueron un importante elemento dentro de la dieta precolombina en muchas áreas. Las vainas de P.vulgaris (frijol común) y las vainas y hojas del P.lunaus (frijol lima) producen fitolitas distintivas con forma de gancho (Bozarth 1986, 1988, 1989b, 1990). Los análisis estadísticos demostraron que muchas de estas fitolitas son significativamente más anchos cerca de la punta que aquellos producidas en otras especies nativas. Las fitolitas del frijol fueron localizadas en un sitio arqueológico del centro de Kansas, basándose en esta característica distintiva (Bozarth 1988, 1989b, 1990). Se han recuperado fitolitas de frijol en cinco sitios de Arizona (Bozarth 1994, 1996).
Además de los anteriores existieron numerosos cultivos, frutas silvestres, nueces, vegetales y tubérculos importantes en la dieta precolombina de Centro América, muchos de los cuales (20 especies) se analizaron fitolíticamente durante la investigación arqueológica en Golfito, Costa Rica (Bozarth 1993b). Las frutas cultivadas y las nueces incluyen Anacardum occidentales (nuez comercial), Ananas comosus (piña), Annona sp (anonillo), Bactris gasipaes (pejibaye), Cariaca papaya (papaya), Cocos nucífera (coco), Inga spp. (guava), Persea americana (aguacate), Psidium guajava (guayaba), y Theobroma cacao (cacao). Las frutas no cultivables comestibles consistían en Anacardium excelsum (nuez silvestre), y Calocarpum corcanum (zapote). La Cresentia alota (jícaro), no comestible, fue procesada para ser utilizada como contenedor o recipiente.
Las especies de algodón fueron domesticadas prehistóricamente en el Nuevo Mundo, Gossypium hirsutum (algodón de montaña) en Mesoamérica y Norteamérica, y G. babadense (algodón de las islas) en Suramérica (Harland 1992). La fibra de G. hirsutum forma ocasionalmente varillas silicificadas huecas y sólidas. Estas fitolitas tienen un rango de longitud de 20 a 540 µm de largo con diámetros de 2-7 µm. Una comparación de estas fitolitas con otros formados en plantas nativas del suroeste americano indica que las varillas huecas, que son curvas o con ramificaciones, son exclusivas del algodón. No obstante, su rareza limita su valor arqueológico (Bozarth 1992 a, 1992 b, 1994). Las fitolitas se producen en las hojas de G. barbadense pero no poseen un valor taxonómico. No se encontraron fitolitas en sus vainas fibrosas (Piperno 1988).
METODOLOGÍA
Tres muestras sedimentarias fueron procesadas para análisis fitolítico, dos del Grupo 18 en Nakbe (18Z.11.16 y 18Z.10.14), y uno del barro negro proveniente de una terraza por el bajo en el lado sur de Nakbe (510A.03.10). Se aislaron las fitolitas en muestras de cinco gramos usando un procedimiento basado en la flotación en líquidos pesados (zinc-bromuro) y la centrifugación (Bozarth 1991). Este procedimiento consiste en seis pasos básicos: (1) remoción de los carbonatos con ácido hidroclórico diluido; (2) remoción de orgánicos coloidales, barros y cienos muy finos por medio de la defloculación con pirofosfatos sódicos, la centrifugación y la decantación a través de filtros de 7 micrones; (3) oxidación de la muestra para remover restos orgánicos; (4) flotación de fitolitas dentro de líquidos pesados desde la fracción más pesada de mineral clástico usando bromuro de zinc concentrado hasta un valor específico de 2.3; (5) lavado y deshidratación de fitolitas con butanol; y (6) almacenaje en seco en frascos de 1-dram. Esta técnica ha sido refinada en el Laboratorio Palinológico de la Universidad de Kansas y ha demostrado ser altamente efectiva al aislar fitolitas de una gran variedad de especies sedimentarias.
Después de la mezcla, una porción representativa de la muestra se montó sobre una platina de microscopio inmersa en aceite, debajo de una cubierta de vidrio de 24 x 40 mm y sellada con esmalte de uñas transparente. Un mínimo de 300 fitolitas de especies económicas fueron examinadas por medio de un microscopio Zeiss con un aumento de 625X. Los resultados se presentan en la Figura 4.
RESULTADOS Y DISCUSIÓN
La meta primordial de este estudio piloto, que fue determinar si las fitolitas se preservan en Nakbe de una forma y una concentración adecuada para su análisis, se logró con la significativa cantidad de fitolitas satisfactoriamente preservadas que fueron recuperadas en las tres muestras estudiadas. Las fitolitas estaban particularmente concentradas en la muestra superior del Grupo 18, que fue una terraza pegada a un complejo residencial de carácter elitista (Figura 5).
El hallazgo taxonómico más relevante fue la identificación de una fitolita de la mazorca de maíz en excelente estado de preservación, de la muestra inferior recolectada en el Grupo 18. Este es un descubrimiento especialmente importante ya que podría ser uno de las primeras fitolitas recobradas en un sitio Maya. Más aún, una fitolita del tipo Lagenaria se identificó dentro de la muestra proveniente de las terrazas cercanas al bajo. Adicionalmente, las fitolitas de palma estaban presentes en las tres muestras, pero esto también podría deberse a la existencia natural de esta planta mucho antes a la ocupación del sitio. La presencia de estatoesporas (cistas esféricas con paredes silíceas producidas en las algas doradas) en las muestras del Grupo 18, podría ser de utilidad para determinar si el sedimento de esa parte del sitio fue transportado desde un bajo en donde se presume la posibilidad del crecimiento de esta alga.
Es necesario hacer un análisis más complejo que incluya un estudio de órganos análogos modernos, antes de establecer el significado cultural que las fitolitas de la palma y las estatoesporas podrían tener. Se observaron fitolitas características tanto de las hierbas como de los árboles en las tres muestras, lo que demuestra que es posible la reconstrucción del paleo-ambiente en este sitio. Ahora que hemos identificado la presencia de fitolitas en los antiguos campos agrícolas en Nakbe, creemos que será posible identificar los cultivos antiguos en el Preclásico, así como la ubicación de sus siembras originales.
En resumen, este estudio demuestra el potencial del análisis fitolítico al tratar de entender las estrategias de subsistencia de los Mayas en el norte de Guatemala. También demuestra que las fitolitas pueden utilizarse para estudiar el impacto ecológico de las culturas precolombinas. El análisis fitolítico sin duda jugará un importante papel en el futuro de las investigaciones arqueológicas en Mesoamérica.
REFERENCIAS
Bozarth, Steven R.
1985 An Analysis of Opal Phytoliths from Rinds of Selected Cucurbitaceae Species. Tesis de Maestría, Department of Anthropology, University of Kansas, Lawrence.
1986 Morphologically Distinctive Phaseolus, Cucurbita, and Helianthus annuus Phytoliths. En Plant Opal Phytolith Analysis in Archaeology and Paleoecology (editado por I. Rovner):56-66. Phytolitharien Occasional Paper No. 1, North Carolina State University, Raleigh.
1987a Phytolith Analysis for Cultigen Identification: 25HN36, 25HN37, and 25HN40. En Prehistoric and Historic Cultural Resources of Selected Sites at Harlan County Lake, Harlan County, Nebraska (editado por M. Adair y K. Brown):469-471. U.S. Army Corps of Engineers, Kansas City.
1987b Diagnostic Opal Phytoliths from Rinds of Selected Cucurbita Species. American Antiquity 52:607-615.
1988 Phytolith Identification of Domesticated Beans (Phaseolus vulgaris) at 14MN328, a Great Bend Aspect Village in Central Kansas. Ponencia presentada al 10th Annual Flint Hills Archaeological Conference. Topeka, Kansas.
1989a Evidence for Zea mays at 14 MN328 Based on Opal Phytolith Analysis. Ponencia presentada al 11th Annual Flint Hills Archaeological Conference, University of Kansas, Lawrence.
1989b Opal Phytoliths. En Final Summary Report, 1986 Archaeological Investigations at 14MN328, a Great Bend Aspect Site Along U.S. Highway 56, Marion, Kansas (por W. Lees, J. Reynolds, T.J. Martin, M. Adair y S. Bozarth):85-90. Kansas Department of Transportation.
1990 Diagnostic Opal Phytoliths from Pods of Selected Varieties of Common Beans (Phaseolus vulgaris). American Antiquity 55:98-104.
1992a Cultigen Phytolith Analysis. En Cultural Dynamics in the Lukachukai Valley: N 13 Project, Draft Report. Statistical Research Technical Project Series 16, Part 2, editado por J.H. Atlschul y S.D. Shelly. Bureau of Indian Affairs.
1992b Fossil Pollen and Phytolith Analysis. En Archaeological Investigations at Lee Canyon, Kayenta Anasazi Farmsteads in the Upper Basin Coconino County, Arizona. Statistical Research Technical Series No.38, pp.135-142. Arizona Department of Transportation.
1993a Maize (Zea mays) Cob Phytoliths from a Central Kansas Great Bend Aspect Archaeological Site. Plains Anthropologist 38 (146): 279-286.
1993b Opal Phytolith Analysis of Shell Middens at Sites P-309B and P313N. En Prehistoric Human Ecology on the Golfo Dulce, Southwestern Costa Rica (editado por J. Hoopes). Department of Anthropology, University of Kansas.
1994 Pollen and Phytolith Analysis. En The Roosevelt Rural Sites Study, Changing Land Use in the Tonto Basin. Statistical Research Technical Series, No.28, U.S. Department of Interior, Bureau of Reclamation.
1996 Pollen and Opal Phytolith Evidence of Prehistoric Agriciulture and Wild Plant Utilization in the Lower Verde River Valley, Arizona. Tesis Doctoral, Department of Geography, University of Kansas, Lawrence.
1997 Pollen and Phytolith Analysis. En State Route 87-Sycamore Creek Archaeological Project, Vol. 2, Draft Report. Statistical Research, Tucson, Arizona.
Brenner, Mark, Barbara Leyden y Michael W. Binford
1990 Recent Sedimentary Histories of Shallow Lakes in the Guatemalan Savannas. Journal of Paleolimnology 4:239-252. Kluwer Academic Publishers, Belgium.
Bush, Mark B., Dolores R. Peperno y Paul A. Collinvaux
1989 A 6000 Year History of Amazonian Maize Cultivation. Nature 340: 303-305.
Castañeda Salguero, César
1997 Sistemas lacustres de Guatemala: recursos que mueren. Editorial Universitaria, Universidad de San Carlos, Guatemala.
Castañeda Salguero, César y César Castañeda Cerna
1994 Evidencias florísticas de la dinámica lacustre (pantanosa) en el área de Nakbe, Petén. En Investigaciones arqueológicas en Nakbe, Petén: resumen de la Temporada de Campo de 1993 (editado por R.D. Hansen):139-151. Informe, Instituto de Antropología e Historia de Guatemala, Monumentos Prehispánicos, Guatemala.
Deevey, E.S., Don S. Rice, Prudence M. Rice, H.H. Vaughan, Mark Brenner y M.S. Flannery
1979 Mayan Urbanism: Impact on a Tropical Karst Environment. Science 206:298-306.
Domínguez Carrasco, María del Rosario
1993 Calakmul, Campeche y su sistema hidraúlico. En Los Investigadores de la Cultura Maya. Publicaciones de la Universidad Autónoma de Campeche, pp.42-46.
Hansen, Richard D.
1994 Las dinámicas culturales y ambientales de los orígenes Mayas: estudios recientes del sitio arqueológico Nakbe. En VII Simposio Arqueológico de Guatemala, 1993 (editado por J.L. Laporte y H.L. Escobedo):115-122. Museo Nacional de Arqueología y Etnología, Guatemala.
1995 Early Environmental Impact: The Ecological Consequences of Incipient Maya Settlement. Informe entregado al National Geographic Society, NGS 4984-93. Washington, D.C.
Jacob, John
1994 Evidencias para cambio ambiental en Nakbe, Guatemala. En VII Simposio Arqueológico de Guatemala, 1993 (editado por J.L. Laporte y H.L. Escobedo):275-280. Museo Nacional de Arqueología y Etnología, Guatemala.
Jones, John
1991 Evaluation of the Potential for Pollen and Phytolith Recovery from Sediments from the Maya Site of Nakbe, Guatemala. Informe en los archivos de PRIANPEG/RAINPEG. Universidad de California, Los Angeles.
Leyden, Barbara
1994 Pollen Analysis from Nakbe, Peten, Guatemala. Informe en los archivos de PRIANPEG/RAINPEG, Universidad de California, Los Angeles.
Mulholland, Susan C.
1987 Phytolith Studies a Big Hidatsa. Tesis Doctoral, Universidad de Minnesota.
Pearsall, Deborah M.
1978 Phytolith Analysis of Archaeological Soils: Evidence for Maize Cultivation in Formative Ecuador. Science 199:177-178. También en Paleoethnobotany: A Handbook of Procedures, Academic Press, San Diego, 1989.
1982 Phytolith Analysis: Application of a New Paleoethnobotanical Technique in Archaeology. American Anthropologist 84:862-871.
Piperno, Dolores R.
1984 A Comparison and Differentiation of Phytoliths from Maize and Wild Grasses: Use of Morphological Criteria. American Antiquity 49:361-383.
1988 Phytolith Analysis: An Archaeological and Geographical Perspective. Academic Press, San Diego.
1990 Aboriginal Agriculture and Land Usage in the Amazon Basin, Ecuador. Journal of Archaeological Science 17:665-677.
1991 The Status of Phytolith Analysis in the American Tropics. Journal of World Prehistory 5 (2):155-191.
Piperno, Dolores R., Karen H. Clary, Richard G. Cooke, Anthony J. Ranere y Doris Weiland
1985 Preceramic Maize in Central Panama: Phytolith and Pollen Evidence. American Anthropologist 87:871-878.
Romero Zetina, Juan J.
1996 Utilización de leña para la elaboración de cal en la península de Tayasal. Proyecto de Investigacion RAINPEG, «Elaboración de Cal de Manera Tradicional por los Mayas de la Zona de Petén, Guatemala». Manuscrito en los archivos de PRIANPEG.
Romero Zetina, Juan J. y Thomas Schreiner
1998 Breve estudio de las zonas ecológicas de Zacatal, Aguada Maya, Nakbe, Petén, Guatemala. Informe en los Archivos de PRIANPEG, Instituto de Antropología e Historia, Guatemala.
Weinstein, Eri
1994 Presence/Absence Analysis of Pollen Recovered from Nakbe, Guatemala: Sediment Samples. En Investigaciones Arqueológicas en Nakbe, Petén: Resumen de la Temporada de Campo de 1993 (editado por R.D. Hansen):346-348. Informe entregado al Instituto de Antropología e Historia de Guatemala.
Wilding, Larry P. y Larry R. Drees
1971 Biogenic Opal in Ohio Soils. Proceedings of the Soil Science Society of America 35:1004-1010.
1974 Contributions of Forest Opal and Associated Crystalline Phases of Fine Silt and Clay Fractions of Soils. Clays and Clay Minerals 22:295-306.
Wilding, Larry P., C.T. Hallmark y N.E. Smeck
1979 Dissolution and Stability of Biogenic Opal. Journal of American Soil Science Society 43:800-802.
Wilding, Larry P., N.E. Smeck y Larry R. Drees
1977 Silica in Soils: Quartz, Cristobalite, Tridymite, and Opal. En Minerals in Soils Environment (editado por J.B. Dixon y S.B. Weed):471-552. Soil Science Society of America, Madison, Wisconsin.
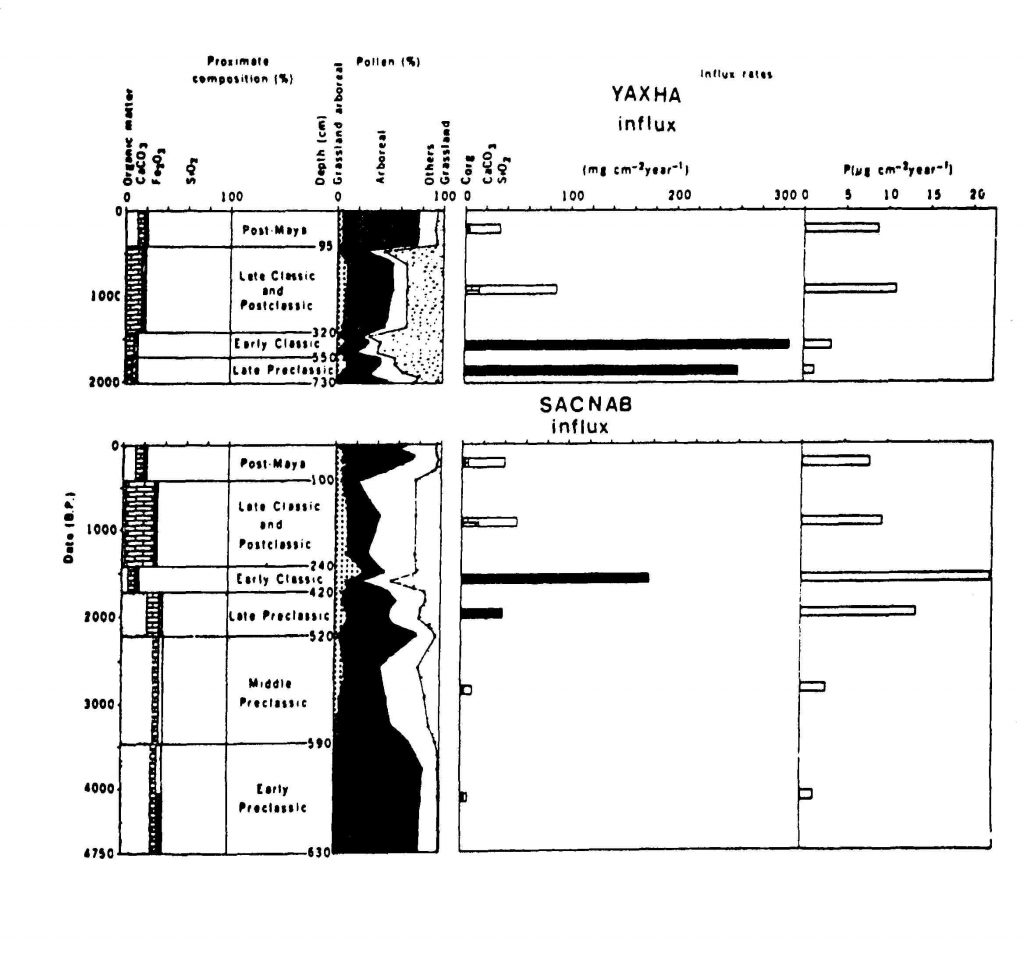
Figura 1 Porcentaje del polen arbóreo y los influjos a los lagos indicados por las muestras extraídas de
los lagos Yaxha y Sacnab. Nótese la reducción de polen arbóreo al final del Preclásico Tardío y el
Clásico Tardío. La mayor cantidad de influjos sucedió al fin del Preclásico Tardío e inicio del Clásico
Temprano (modificada de Deevey et al. 1979: Figura 4)
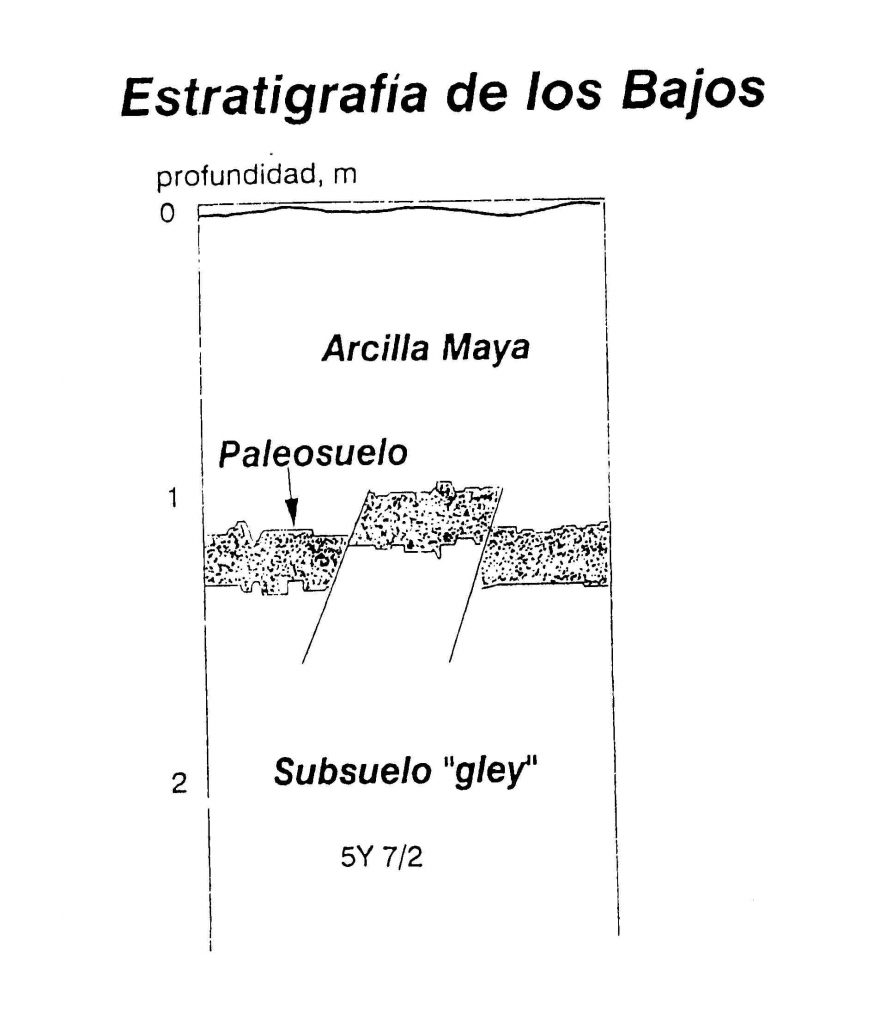
Figura 2 Perfil típico de las capas estratigráficas de los bajos alrededor de Nakbe. Las trincheras y pozos
revelaron una capa de paleo-suelo enterrada debajo de sedimentos de arcilla Maya (modificada de
Jacob 1994:279)
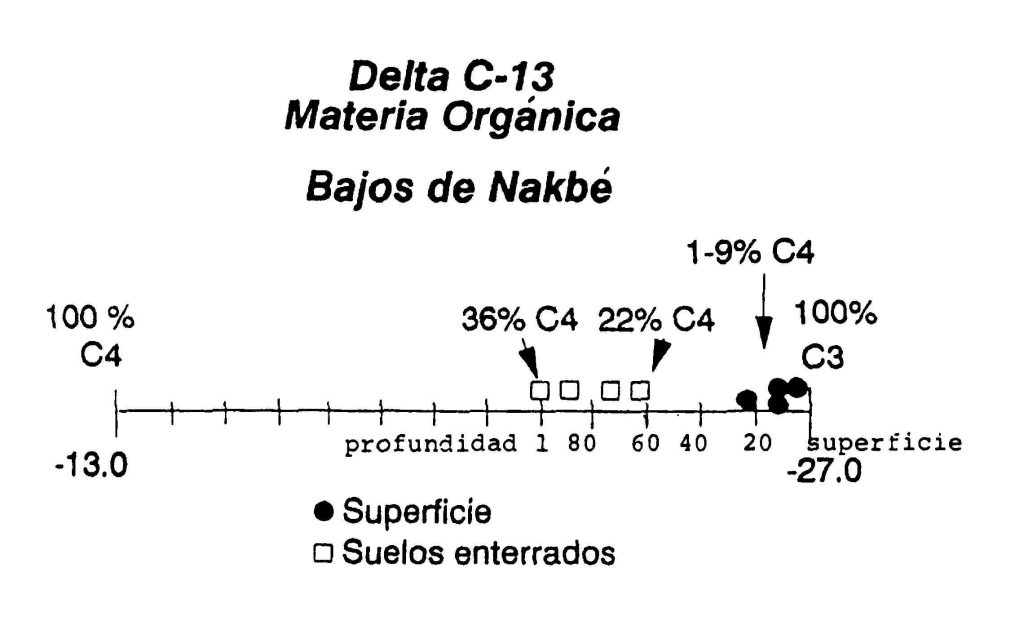
Figura 3 La composición isotópica de los perfiles estratigráficos de los bajos indica que la capa de paleo-
suelos consistió de plantas del tipo C4 (maíz y zacate), mientras la capa de sedimento consistió de
plantas C-3 (el bosque actual; modificada de Jacob 1994:280)
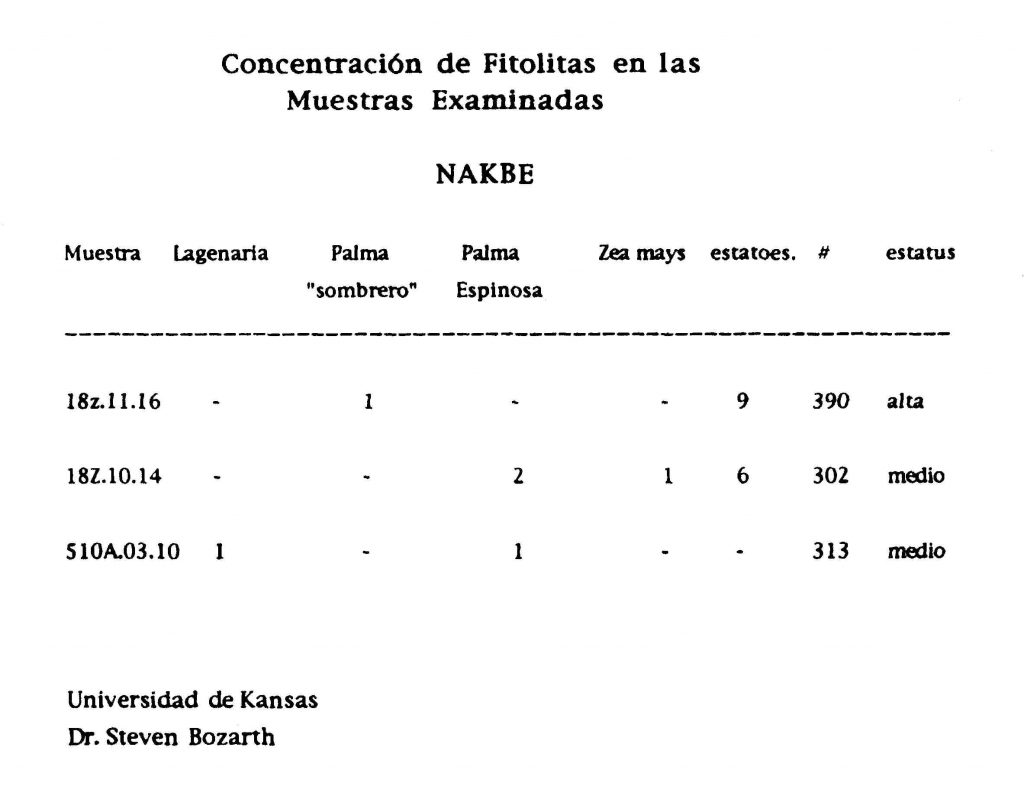
Figura 4 Resultados de un mínimo de 300 fitolitas de especies económicas examinadas con un
microscopio Zeiss con un aumento de hasta 625X (Bozarth 1997:15)
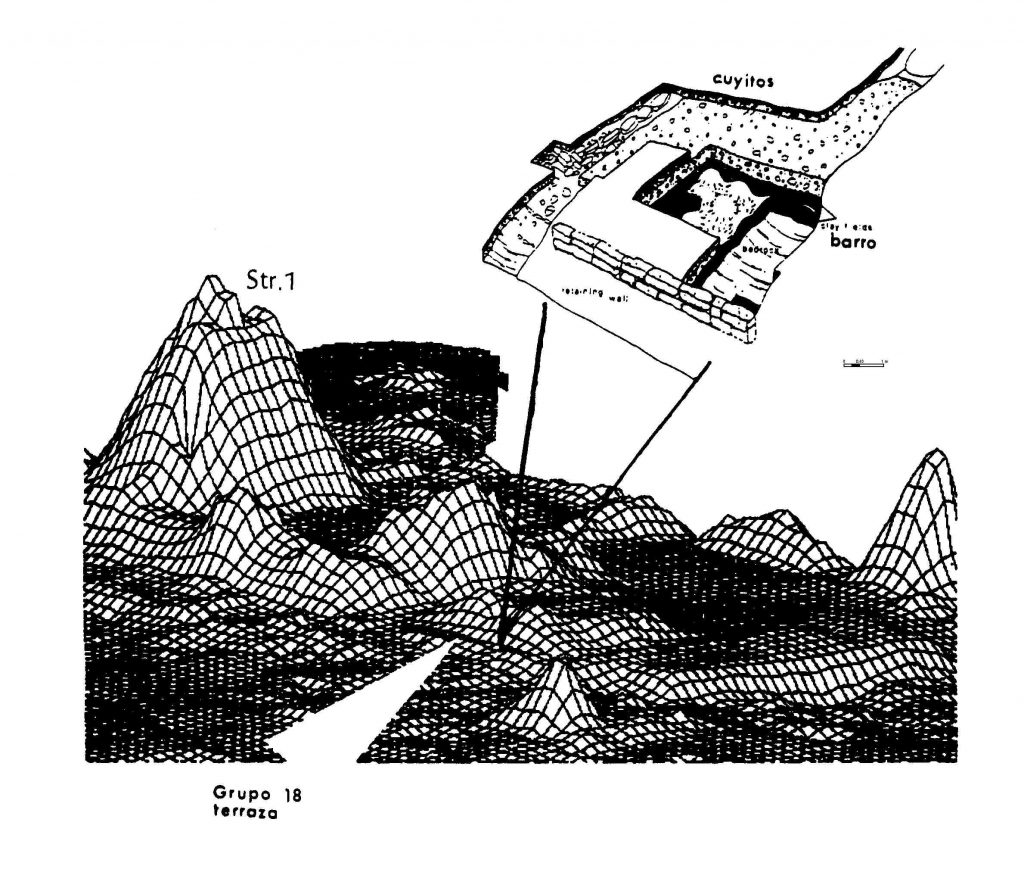
Figura 5 Grupo 18. Una terraza construida de barro importado se encuentra pegada al lado sur del
complejo residencial de carácter elitista en el Grupo Oeste, Nakbe