

Descargar este artículo en formato PDF
Götz, Christopher M.
2008 Manjares del pasado: Contraste del aprovechamiento faunístico entre sitios prehispánicos costeros y de tierra adentro de las Tierras Bajas del Norte. En XXI Simposio de Investigaciones Arqueológicas en Guatemala, 2007 (editado por J. P. Laporte, B. Arroyo y H. Mejía), pp.964-983. Museo Nacional de Arqueología y Etnología, Guatemala (versión digital).
62
MANJARES DEL PASADO: CONTRASTE DEL APROVECHAMIENTO FAUNÍSTICO ENTRE SITIOS PREHISPÁNICOS COSTEROS Y DE TIERRA ADENTRO DE LAS TIERRAS BAJAS DEL NORTE
Christopher M. Götz
Facultad de Ciencias Antropológicas, Universidad Autónoma de Yucatán
Palabras clave
Arqueología Maya, Yucatán, Campeche, Zooarqueología, dieta Maya, uso de animales, consumo, Clásico, Postclásico
Abstract
DELICACIES OF THE PAST: CONTRASTING THE EXPLOITATION OF FAUNA BETWEEN PREHISPANIC COASTAL AND INLAND SITES OF THE NORTHERN LOWLANDS
This work will compare systems of vertebrate animal exploitation between coastal and inland sites of the northern lowlands of the Maya area. Zooarchaeological data will be presented on the use of animals at Classic as well as Postclassic sites of the region under investigation, indicating the fundamental differences between coastal and inland exploitation and use of vertebrate animals and demonstrating the primary, commercial, consumer, and environmental particulars in Prehispanic times. The sites to be studied are located throughout the northern zone of the Yucatan peninsula, including Xcambo, Siho, Chichen Itza, and Dzibilchaltun in Yucatan, Mexico, and Champoton in Campeche, Mexico.
El presente artículo tiene como objetivo describir e investigar, desde un enfoque zooarqueológico, las diferencias a nivel del aprovechamiento de animales vertebrados entre asentamientos de dos ecosistemas: el costero y el de tierra adentro, de las Tierras Bajas del Norte del área Maya. Un énfasis del trabajo es la comparación de los perfiles taxonómicos resultantes del material faunístico excavado en cinco sitios distribuidos sobre el área mencionada, para revelar, tras un análisis zooarqueológico y contextual detallado, patrones de uso que puedan formularse en cuanto a modos generales o específicos del aprovechamiento de animales.
El enfoque zooarqueológico de esta comparación se refiere a los restos esqueléticos de animales que habitaban un conjunto en tiempos pasados, pero trata a estos en particular, referente a la información antropológica que puedan aportar, investigando y, en el mejor de los casos, reconstruyendo así la interacción entre humanos y animales en el pasado.
Partiendo del título del trabajo, se quiere someter a la investigación únicamente los restos de animales que presuntamente sirvieron para el consumo, por lo que se hará la referencia solamente a materiales faunísticos procedentes de basureros asociados a estructuras domésticas, no se abarcará el uso ritual de los animales en este tratado. Sin embargo, los perfiles taxonómicos de los basureros domésticos sí pueden informar, como ya se ha demostrado repetidamente (O’Connor 2003), sobre la obtención primaria de los animales, el comercio y el consumo que presuntamente sucedieron en tiempos prehispánicos, combinándolos con datos óseos como por ejemplo la frecuencia esquelética o el análisis de las marcas antrópicas en los huesos.
Importantes estudios en las últimas décadas, se han dedicado a la alimentación de los Mayas prehispánicos, ya sea basándose en las fuentes etnohistóricas, en análisis antropológico-físicos o en análisis zooarqueológicos (Márquez 1991). Se cree pertinente, sin embargo, tratar el tema de la alimentación Maya prehispánica comparando de manera muy particular acumulaciones de asociación contextual y naturaleza semejante, arrojando así nuevos datos, resultado de trabajos recientes. Los datos que se presentan en este trabajo no son por lo tanto logro de una repetición y reformulación de los datos antiguos, sino parten de una reciente investigación comparativa a gran escala, llevada a cabo en las Tierras Bajas Mayas del Norte (Götz 2007).
Al final de la presente versión digital se ha incluido una serie de esquemas y fotografías que ilustran los planteamientos expuestos en este trabajo (Figuras 9 y 10).
MATERIALES Y TÉCNICAS
Los dos ecosistemas que en este estudio se sujetan a comparación, a través de los sitios arqueológicos y los materiales óseos faunísticos de estos, se sitúan en el ámbito de las Tierras Bajas del Norte, entre la zona de bosques bajos y caducifolios (Carnevali, Ramírez y González-Iturbe 2003:55) y la costa del Golfo de México, abarcando político-geográficamente a los estados modernos mexicanos de Campeche y Yucatán. El material faunístico de los sitios abarca entre el Clásico Tardío y Postclásico Tardío, resumiendo así una muestra de información zooarqueológica de aproximadamente 700 años. Es importante recalcar que los basureros de los sitios cotejados parecen corresponder, por la asociación arquitectónica en todos los casos, a formaciones de grupos dominantes de los sitios. Los asentamientos prehispánicos costeros sujetados a la comparación son Xcambo y Champoton, los de tierra adentro incluyen a Chichen Itza, Siho y Dzibilchaltun, caracterizados e importantes en este trabajo por los puntos mencionados a continuación (Figura 1).
El sitio de Xcambo, Yucatán, está ubicado cerca de la costa del Golfo de México, en un área de manglares y petenes de suelos enriquecidos por la sal de la cercana agua marina. La ocupación más importante de Xcambo oscilaba entre el Clásico Temprano y Clásico Tardío (250-800 DC), fecha asignada también a la muestra de 1500 especímenes esqueléticos de animales de Xcambo, procedente de rellenos constructivos de plataformas y patios. Una de las principales funciones del lugar aparenta haber sido el comercio de bienes como la sal (Andrews 1997; Sierra 1999; Sierra y Martínez 2001).
El sitio de Champoton se localiza directamente en la costa campechana del Golfo de México, a orillas de la boca del río Champoton, por lo que se asigna también a Champoton una posición importante en el comercio de larga distancia. El sitio estuvo ocupado desde el Preclásico, pero el material faunístico analizado, 1720 especímenes de un gran basurero asociado al Grupo 5, procede de la zona postclásica del sitio (1200–1500 DC; Folan et al. 2004; Forsyth y Jordan 2005).
Chichen Itza, ciudad cosmopolita Maya del Clásico Terminal y Postclásico Temprano (1050-1200 DC), se encuentra en el oriente del estado yucateco (Schmidt 2005), arrojó huesos faunísticos de un gran basurero hallado en un grupo habitacional de alto estatus de la Serie Inicial, fechado para el Clásico Terminal y Postclásico Temprano. Los huesos faunísticos del basurero, 4002 en su totalidad, se localizaron en conjunto con más de 300,000 tiestos cerámicos, restos de artefactos pétreos y algunos huesos humanos (Schmidt 2003, 2005).
Del sitio de Dzibilchaltun, situado en la planicie norte a unos 50 km de la costa, procede un conjunto faunístico, fechado para el Clásico Tardío y Terminal, excavado inmediatamente al norte de la Plaza Sur e igualmente asignado al grupo socioeconómicamente dominante del sitio. Los 1125 especímenes faunísticos del basurero se ubicaron en conjunto con más de 11,000 tiestos cerámicos, restos de artefactos líticos y conchas (Maldonado, comunicación personal; Andrews y Andrews 1980).
Por último, Siho, del occidente del estado yucateco, representa un asentamiento de ocupación del Clásico Tardío y Terminal, del cual se obtuvieron 100 especímenes faunísticos de un basurero asociado al grupo habitacional central, de estrato socioeconómico elevado y posiblemente el más importante del lugar (Cobos et al. 2002, 2004).
Respecto al análisis zooarqueológico seguido en esta investigación, puede mencionarse que abarcó, de manera estandarizada en la zooarqueología, la obtención de los datos primarios y el cálculo de algunos datos secundarios (Reitz y Wing 1999:142-238). Los datos primarios consistieron en la identificación taxonómica, osteológica y tafonómica de los materiales faunísticos. Estas se efectuaron mediante el uso de una colección comparativa de vertebrados modernos, una colección de fotografías digitales de especímenes esqueléticos de la región Maya (EA.FLMNH 2003) y manuales de osteología faunística de la región tratada. La terminología taxonómica de los especímenes proviene de recientes publicaciones sobre la fauna regional, mientras que el análisis tafonómico de los huesos se rigió por pertinentes trabajos acerca del tema (Lyman 2000; Reixach 1986; White 1992).
Los datos secundarios, manipulaciones matemáticas de los datos primarios, incluyeron las cuantificaciones de los especímenes expresadas en el número de ejemplares identificados por taxon (NISP, del término inglés «Number of Identified Specimens»; Lyman 2000:511), y el número mínimo de individuos (MNI, del término inglés «Minimum Number of Individuals», que se refiere a la cantidad de individuos de un taxon y contexto, reconstruidos con base en la frecuencia esquelética y el NISP; Lyman 2000:510), así como la distribución de fragmentos por porción corporal de cada taxon en los contextos individuales (frecuencia esquelética). Todos estos cálculos forman la base de las comparaciones estadísticas usadas a lo largo del trabajo (Reitz y Wing 1999). Para las comparaciones entre los contextos se utilizó el número de especímenes identificados por taxon (NISP) y el número mínimo de individuos (MNI).
RESULTADOS
El material de Xcambo proviene de rellenos constructivos y por eso no representa tan claramente un basurero acumulado, como en el caso de los demás sitios discutidos aquí. Sin embargo, se valora el material de Xcambo como material de desecho, porque es muy probable que no se efectuó una clara formación de basureros en este sitio, usándose el material por parte de los antiguos habitantes más bien –dictado por la ubicación del sitio en una isla- para fines constructivos. En general, el material faunístico de Xcambo marca una gran diversidad taxonómica, se encontraron una serie de especies de animales de todas las clases de vertebrados, desde aves, mamíferos, peces y reptiles. Entre las aves se localizaron huesos de pavos de monte (Meleagris ocellata), pelícano (Pelecanus sp.), bobo (Sula sp.), garza (Ardea herodias y Egretta sp.), pato, (Anatidae), chorlito (Batramia sp.), gallinola (Gallinula cf. chloropus) y amazona (Amazona sp.). Mamíferos hubo ejemplares tanto de especies terrestres como acuáticas, como zarigüeya (Philader opossum), armadillo (Dasypus novemcinctus), aguti (Agouti paca), conejo de monte (Sylvilagus sp.), perro (Canis lupus familiaris), zorra gris (Urocyon ciniereoargenetus), mapache (Procyon lotor), puma (Puma concolor), manatí (Trichechus manatus), tapir (Tapirus bairdii), pecarí de collar (Pecari tajacu), venado temazate (Mazama sp.), venado cola blanca (Odocoileus virginianus), delfín (Dephinidae) y foca (Monachus tropicalis). La igualmente variada ictiofauna de Xcambo contiene especímenes del tiburón gato (Ginglymostoma cirratum), mako (Carcharhinus sp.), tigre (Galeocerdo cuvieri), cazón (Rhizoprionidon terranpvae) y martillo (Sphyrna cf. tiburo), así como rayas (Dasyatis sp. y Aetobatus sp.) y osteíctios, como bagre (Ariopsis felis), pez diablo (Opsanus sp.), robalo (Centropomus sp.), mero (Epinephelus sp.), ulúa (Carangidae), pargo (Lutjanus sp.), mojarras marinas (Calamus leucostomus) y gurrubata (Micropogonias sp.). Los reptiles de Xcambo, finalmente, se dividen entre huesos de cocodrilo (Crocodylus cf. moreletti), las especies de tortuga caguama (Caretta caretta), verde (Chelonia mydas), carey (Eretmochelys imbricata), blanca de río (Dermatemys mawii), casquito (Kinosternon sp.), terrestre (Terrapene sp.) y jicotea (Trachemys sp.), así como iguana negra (Ctenosaura cf. Similis; Figuras 2 y 3).
Los huesos arqueológicos faunísticos de Champoton son taxonómicamente menos variados. La gran mayoría de los huesos del basurero son de aves, entre las cuales aparecen predominantemente pavos, tanto del monte como doméstico (Meleagris gallopavo). Además se ubicaron huesos de garza, gallareta (Fulica cf. americana) y chachalaca (Ortalis vetula). Los restos de mamíferos están porcentualmente inclinados hacia un taxon, mostrándose mucho más restos de perro que en los otros contextos. También hubo huesos de venado cola blanca, en segundo lugar en frecuencia, temazate, conejo de monte, comadreja, jaguar (Panthera onca), pecarí de collar, manatí y tapir. Entre la gran cantidad de huesos de tortugas de mar destacan restos de la tortuga verde, carey y caguama, además de la tortuga blanca de río, pero también tortuga guau (Staurotypus triporcatus), casquito y jicotea, así como iguana negra, serpiente boa (Boa constrictor) y cocodrilo de pantano. La muestra íctia está dominada por huesos del bagre, robalo y cazón, pero se encontraron también huesos de pez diablo, aguja (Belonidae), mero, mojarra, pargo y gurrubata (Figuras 2 y 3).
Los huesos faunísticos del basurero de la Serie Inicial de Chichen Itza se dividen en mamíferos (94%), seguido por reptiles (4%), aves (2%) y peces óseos (2%). Entre los huesos de mamíferos se localizaron especímenes de conejo de monte, aguti, perro doméstico, pisote (Nasua narica), armadillo, zarigüeya, venado temazate, venado cola blanca, pecarí de collar y pecarí de labios blancos (Tayassu pecari). Entre los reptiles habían mayoritariamente, con un 95%, huesos de iguanas negras, tortugas jicotea y casquito. Los huesos de aves pertenecen a las especies de hocofaisán (Crax rubra), pavo del monte y pavo sin especificar (Meleagris sp.), probablemente también pavo del monte. Los huesos de osteíctios no pudieron determinarse más allá del nivel de clase debido a una fuerte erosión (Figuras 2 y 3).
El perfil taxonómico de Dzibilchaltun marca las especies de venado cola blanca, venado temazate, pecarí de collar, conejo de monte, zarigüeya (Didelphis sp.), comadreja (Mustelidae) y perro doméstico, mientras que entre las aves se manifestaron huesos de chachalaca, tórtola (Columbia) y pavo, tanto del monte como posiblemente doméstico. Los reptiles y anfibios, finalmente, están comprendidos por restos de la tortuga caguama y de sapo (Bufo marinus), mientras que el material íctio únicamente reveló la existencia de bagre marino (véase Figura 2 y 3).
El material faunístico de Siho, dividido entre el Clásico Tardío (93%) y Terminal (7%) según la asociación cerámica, comprende las huesos de venado cola blanca, venado temazate, tortuga terrestre (Terrapene sp.), iguana negra, serpiente boa (Boa constrictor) y cascabel (Crotalus sp.), raya (Rajiformes) y bagre marino (Figuras 2 y 3).
DISCUSIÓN
Los perfiles taxonómicos descritos en los párrafos anteriores reflejan de manera obvia una fuerte diferencia entre los sitios costeros y los sitios tierra adentro en cuanto a la presencia arqueológica de especies de animales. Se observó que no solamente varía la diversidad, sino también el carácter imponente y los porcentajes de algunas especies en los conjuntos óseos.
Antes de entrar, sin embargo, a una comparación de los perfiles hay que revisar y argumentar porqué las acumulaciones óseas de animales informa sobre el uso de estos en el pasado, lo que se refiere en principio al análisis tafonómico de las muestras. Claro está, los animales no solamente constituyen el ambiente prehispánico, sino ocuparon y ocupan también las zonas de los sitios arqueológicos después de su abandono. Es por eso que la asignación de un origen antrópico a una simple acumulación ósea, a pesar de que esta se haya ubicado en un sitio arqueológico, representa frecuentemente retos analíticos e interpretativos. Cabe señalar que en el ámbito de las culturas precolombinas no hubo gran cantidad de animales domesticados (Valadez 2003), cuya simple presencia en un conjunto asignaría un origen humano. Fuera de esto también se manifiesta una general pobreza de marcas resultantes de actividades humanas de preparación de los cuerpos de animales en contextos mesoamericanos, presumiblemente debido a la industria lítica usada o técnicas de preparación de otra índole, por lo que frecuentemente ni las huellas superficiales ni la presencia de animales específicos pueden argumentar convincentemente el origen humano de restos óseos faunísticos.
En relación a los cinco conjuntos óseos discutidos se considera sin embargo, que se puede declarar una asociación con antiguas actividades humanas para la mayor parte del material, principalmente por tres razones. En principio se parte del hecho que los restos estaban asociados a materiales culturales en grandes cantidades, mezclados con tiestos y otros artefactos en capas estratigráficamente definidas. Estas capas, de huesos de animales y materiales culturales, se ubicaban además al interior de sitios arqueológicos concretos, por lo que al menos la formación del contexto óseo puede considerarse de origen prehispánico. El origen prehispánico de los huesos o, dicho de otra manera, identificar a los pobladores prehispánicos como los causantes del deceso de los animales está anclado a este primer punto, pero se basa también en el hecho de que al menos una parte de los especímenes óseos, de la mayoría de las especies representadas, está provista de al menos algunas marcas de posible actividad humana, como cortes, fracturas del hueso en estado fresco, indicios de haber sido hervido, quemado, etc. Desde que estas huellas -con excepción de la última mencionada, que sí presenta un problema de evaluación cuando aparece sin las demás en un conjunto (Lyman 2000:384ss.; White 1992:335, 344)- se han podido reproducir en experimentos arqueológicos (Fisher 1995; Pumajero 1990; Reixach 1986), queda no solamente afirmada su asociación con actividades humanas, sino también explicado por qué habría una relativa pobreza de huellas en el material óseo faunístico.
Por último, hay que considerar que el volumen de acumulación, si bien aún no se puede especificar exactamente en cuánto tiempo se formó, si parece demostrar igualmente que los humanos fueron los responsables, ya que la ubicación de los conjuntos en los sitios y a nivel de la geografía peninsular no hace posible que se trate de conjuntos formados por alguna catástrofe o algo semejante. Claro está, no se puede afirmar que todos los huesos -y con esto todas las especies presentes- hayan sido producto de las antiguas actividades de obtención y preparación humanas. En específico los animales de talla menor y poco representados en los conjuntos podrían ser fruto de algún proceso natural, pero la gran mayoría, así como el perfil taxonómico general -y mucho más allá de esta afirmación no se puede llegar- parece ser de origen prehispánico.
Uniendo la información de los perfiles taxonómicos en general se observa el siguiente patrón (Figuras 4 a 8). Empezando con los sitios costeros de Xcambo y Champoton se muestra que la fauna encontrada en los basureros y presumiblemente aprovechada por los Mayas prehispánicos, es variada pero enfatiza los animales procedentes del área lacustre, semejante a otros contextos costeros ya investigados (Márquez 1991). Una gran variedad de peces, aves, reptiles y mamíferos está presente, pero se nota, al considerar las cuantificaciones del material óseo (el NISP y el MNI), que en ambos sitios costeros hay un acentuado énfasis en ciertos animales, mismo que caracteriza en particular a los sitios y los difiere tanto entre sí, como de los sitios tierra adentro. Entre los mamíferos hay una presencia acentuada –pero porcentualmente mucho menos relevante que en sitios tierra adentro- de perros y venados cola blanca. La avifauna arqueológica costera está dominada, más en Champoton pero en proporción a la cantidad total de restos de ave también en Xcambo, por pavos, viéndose en Champoton inclusive las dos especies, el pavo silvestre y el pavo doméstico. La herpetofauna de los sitios costeros se caracteriza por una abundancia de tortugas, donde, en cierta forma esperadamente, dominan las tortugas marinas y de fangos, animales entonces de ambientes cercanos a los sitios en cuestión (Lee 2000).
Como era de esperarse, la ictiofauna de Xcambo y Champoton proviene de las aguas del golfo de México, ya que en el material identificado no se halló ninguna especie de pez óseo o cartilaginoso restringida a ambientes distintos, como serían por ejemplo aguas dulces o el mar Caribe (Robins, Ray y Douglass 1986). Las especies de peces identificadas en los contextos parecen haberse capturado en la cercanía a las orillas, ya que al menos las especies claramente identificadas ocurren principalmente en las zonas litorales (Robins, Ray y Douglass 1986). El énfasis en los litorales se manifiesta al parecer ––aunque admitidamente partiendo de un cálculo de fragmentos identificados, ya que establecer un MNI para peces resulta muy difícil–– en la ocurrencia acentuada, tanto en Champoton como en Xcambo, de las especies de robalo y bagre, ambas habitando aguas cercanas de la costa e inclusive en estuarios (Robins, Ray y Douglass 1986:83, 128ss.).
El conjunto de los perfiles taxonómicos de los sitios tierra adentro indica que los habitantes prehispánicos aprovechaban una variedad de fauna de las zonas adyacentes a sus asentamientos. La fauna reflejada en los perfiles contiene desde los más grandes animales existentes en el lugar a unas especies de tamaño muy reducido, abarcando en este sentido las clases vertebradas de mamíferos, aves y reptiles. Sin embargo, también se nota que en los sitios tierra adentro hay un énfasis en ciertos animales, reconocible tanto en la cantidad de fragmentos como en el MNI, específicamente en venados cola blanca, pavos de monte e iguanas negras. La abundancia de estos animales en los contextos parece manifestar una especialización en la caza y en el consumo. La mayoría de los animales de los tres basureros habita bosques secundarios, concordante con la imagen del patrón de asentamiento bosquejada para el Clásico Maya y para las técnicas agrícolas de la milpa. Pero son las especies arriba mencionadas que abundan –mucho más que otras- en aquellos bosques secundarios, zonas milperas e incluso cerca de asentamientos humanos (Leopold 2000:313, 577-8; Lee 2000:192), –o sea en las zonas adyacentes a los asentamientos prehispánicos durante su ocupación- y que estarían más que otros animales al alcance de los cazadores.
Al considerar los perfiles taxonómicos de los cinco contextos comparados en términos de las ecozonas de procedencia se ve, a pesar de las notables diferencias entre la variedad de fauna encontrada por sitio, una semejanza en cuanto a los modos de obtención, partiendo aquí obviamente de la suposición antes argumentada de que los animales de los basureros sean en su gran mayoría producto de la acumulación humana. Se observa que tanto en los sitios costeros como en los sitios tierra adentro se siguió una obtención, o sea cacería, oportunista de una gran variedad taxonómica de animales de los respectivos ecosistemas. Animales de distintas clases taxonómicas y de distinto tamaño formaron parte de lo que económicamente fue aprovechado por los Mayas de las Tierras Bajas del Norte, pero entre la variedad hay, tanto en las zonas costeras como en la región tierra adentro, un acento en algunas pocas, mismas que argumentan a favor de preferencias alimenticias como de ecosistemas a aprovechar para la caza.
Es así que en la costa se aprovechaban porcentualmente más tortugas, en especial marinas, perros, aves y peces de estuarios (bagres y robalos), mientras que la fauna más escogida en los sitios tierra adentro comprende a venados cola blanca, pavos del monte e iguanas negras. El perfil de aprovechamiento de estas especies en las Tierras Bajas del Norte se muestra en los sitios costeros y tierra adentro, aparentemente sin importar la cronología del conjunto, y se nota igualmente que el registro iconográfico -específicamente el códice Madrid- (Lee 1985) y las fuentes etnohistóricas de tiempos coloniales (De la Garza 1983; De Landa 2003) afirman el beneficio sobre todo de estos animales para tiempos del Postclásico Tardío y la época Colonial temprana.
Podría argumentarse que el “énfasis en ciertas especies” se deba a condiciones tafonómicas de destrucción y preservación diferencial, donde aquellos animales vertebrados que hoy se ven como los más importantes, sean en realidad los de un esqueleto más robusto y con mejores condiciones de preservarse en relación a otros, o bien, que aquellas especies de poca presencia hayan sido frecuentes pero sus restos no se guardaron adecuadamente o fueron usados para otros fines, formando contextos arqueológicos aún no ubicados. En el caso de los sitios costeros, este argumento tendrá que ser analizado seriamente, porque precisamente los muy frecuentes huesos de tortugas marinas, son muy gruesos y robustos, mientras que los huesos de peces poseen una consistencia contraria. Sin embargo, se tiene una gran cantidad de huesos de pescados, mostrando la buena preservación del conjunto, mientras que del gran número de huesos de tortugas marinas no se pudo, -debido a la naturaleza poco diagnóstica de muchos fragmentos-, calcular valores altos del MNI, por lo que, contrario al ‘argumento tafonómico’ expuesto anteriormente, parece indicar que al menos el MNI de las tortugas marinas inclusive sea demasiado bajo en referencia a los restos de estas (O’Connor 2000:54ss.). En sitios tierra adentro son también los venados cola blanca animales de una estructura ósea robusta, pero también los animales poco representados poseen huesos fuertes y los contextos tuvieron igualmente una acentuada presencia de huesos de animales menores, argumentándose entonces que la presencia acentuada o no de las especies, al menos no parece deberse a condiciones de preservación diferencial en el contexto.
Fuera de los animales comunes en los contextos y del probable patrón de aprovechamiento común que se observa reflejado en los contextos discutidos, hay que hacer énfasis en un punto más, en la existencia de los animales poco comunes en los conjuntos arqueofaunísticos. El primero, pero aquí únicamente hipotético punto se refiere a una dicotomía de la presencia, poca, pero sin duda relevante, de animales tierra adentro en los sitios costeros y animales costeros en sitios tierra adentro. Se ve que en el sitio de Siho, Dzibilchaltun y Chichen Itza se encontraron huesos de peces, marinos todos estos, identificados como bagre marino y raya, mientras que en Dzibilchaltun se ubicaron además huesos de tortuga marina. Por el otro lado hubo, tanto en Xcambo como en Champoton, una presencia reducida pero innegable de animales de bosques secundarios de tierra adentro, además de la fauna marina y lacustre está presente.
Es tentador interpretar esta ocurrencia como un reflejo de la dieta Maya de élite (Emery 2003), que hacía necesario traer por ejemplo peces de las costas a sitios tierra adentro como lo propone Carr (1986) o animales de bosques secundarios a los sitios costeros. A pesar de que los restos de peces de este análisis son partes que pudieron servir también como artefactos, como las agujas de raya y bagre, se conocen otros ejemplos de restos craneales y postcraneales de sitios como Dzibilchaltun (Wing y Steadman 1980), Mayapan (Pollock y Ray 1957) o Chichen Itza (Carr 1986), donde se han hallado restos de peces marinos en dichos sitios tierra adentro. Los animales de tierra localizados en los sitios costeros son en su totalidad especies que habitan el bosque secundario y que en su gran mayoría tienen requerimientos ecológicos versátiles, lo que vale por ejemplo para los pavos, pecarís y venados. De estas especies también hay, fuera de los sitios aquí discutidos, ejemplos de otras investigaciones. En sitios aún más exclusivamente ‘costeros’ como aquellos ubicados en islas se han encontrado restos de estas especies, como se observa en el caso de Cozumel (Hamblin 1984:122ss.) o Isla Cerritos (Carr 1987). Se afirma entonces que probablemente hubo al menos un limitado movimiento de especies de las costas hacia tierra adentro –peces y tortugas enteros o por partes en este caso- pero se hace difícil argumentar a favor de un movimiento revés, de especies tierra adentro hacia la costa.
Un caso ciertamente particular, pero que aún así no rompe con el esquema de obtención descrito arriba, es el hallazgo de restos de foca en Xcambo. Los fragmentos craneales y de las extremidades indican la presencia de al menos dos individuos que fueron probablemente cazados ya sea en la playa o en las aguas poco profundas de la costa y arrastrados al sitio, donde se procesaron, quizá para consumo.
Finalmente debe discutirse también la aparición de huesos de una especie al parecer verdaderamente foránea al área Maya, el pavo doméstico, cuyos restos se localizaron en las acumulaciones postclásicas de Champoton. El pavo doméstico en Mesoamérica corresponde hoy día a la especie Meleagris gallopavo, cuya área de expansión, según los ecosistemas actuales, se limita a la región entre el centro de México y al sur de EE.UU (Howell y Webb 2004:225). Del centro de México se tienen hallazgos de esta especie, morfológicamente distinta al pavo endémico de la zona Maya, el Meleagris ocellata (Steadman 1980), desde el Preclásico, pero los conjuntos faunísticos del área Maya han arrojado hasta la fecha únicamente contextos del Postclásico con esta especie (Hamblin 1984; Götz 2007), ubicados tanto en la isla de Cozumel, como en el ya mencionado Champoton. El pavo doméstico parece haber sido entonces un animal verdaderamente llegado al área Maya durante el final de la época prehispánica, distribuyéndose en las zonas costeras, pero que no ingresó a las regiones de tierra adentro, hasta la venida de los españoles.
CONCLUSIÓN
En resumen se puede manifestar que los modos de aprovechamiento de recursos animales en tiempos prehispánicos en las Tierras Bajas del Norte estuvieron regidos por las diferencias de ecosistemas en las que estaban los respectivos sitios, pero dentro de los ecosistemas se siguió un patrón semejante en cuanto a la concentración en unas especies, ciertamente diferentes entre costa y tierra adentro, pero también en el aprovechamiento oportunista en general. Esta suposición, por tan lógica que parezca, indica también que los patrones alimenticios de los Mayas prehispánicos, aquí tratados frente a la o las clases socioeconómicamente dominantes, no seguía un patrón uniforme, como aquel que se puede por ejemplo observar en las manifestaciones artefactuales de la misma cultura y área.
AGRADECIMIENTOS
El presente trabajo ha sido posible con el apoyo de varios directores de los proyectos arqueológicos que a continuación se mencionan: se agradece al Dr. Peter J. Schmidt (Proyecto Arqueológico Chichen Itza, CRY INAH), al Mtro. Rubén Maldonado (Proyecto Arqueológico Dzibilchaltun, CRY INAH), a la Dra. Telma Sierra Sosa (Proyecto Arqueológico Xcambo, CRY INAH), al Dr. Rafael Cobos (Proyecto Arqueológico Siho, FCA UADY) y al Dr. William Folan (Proyecto Arqueológico Champoton, CIHS UAC), por haber proporcionado el material óseo faunístico para su análisis.
REFERENCIAS
Andrews, Anthony P.
1997 La sal entre los antiguos mayas. Arqueología Mexicana 5 (28):38-45. México.
Andrews, E. Wyllys IV y E. Wyllys Andrews V
1980 Excavations at Dzibilchaltun, Yucatan, México. Middle American Research Institute, No.48. New Orleans.
Carnevali, Germán, Iván M. Ramírez y José Antonio González-Iturbe
2003 Flora y vegetación de la Península de Yucatán. En Naturaleza y Sociedad en el Área Maya (editado por P. Colunga-Garcían y A. Larqué), pp.53-68. Centro de Investigaciones Científicas de Yucatán. México.
Carr, Helen Sorayya
1986 Preliminary Results of Nonmolluscan Faunal Analysis at Chichen Itza. (Informe no publicado).
1987 Preliminary Analysis of Nonmolluscan Faunal Remains from Isla Cerritos, Yucatan. (Informe no publicado).
Cobos, Rafael, Vera Tiesler, Pilar Zabala, Lilia Fernández, Armando Inurreta, Nancy Peniche May, Ma. Luisa Vázquez y Diana Pozuelo
2002 El Surgimiento de la Civilización en el Occidente de Yucatán: los Orígenes de la Complejidad Social en Sihó. Informe de Actividades de la Temporada de Campo 2001. Universidad Autónoma de Yucatán, Facultad de Ciencias Antropológicas, Mérida, Yucatán.
Cobos, Rafael, Nancy Peniche May, Edgar D. Pat Cruz, Socorro Jiménez Álvarez, Lilia Fernández, Vera Tiesler Blos, Christopher M. Götz y Alfonso Lacadena
2004 Proyecto Arqueológico El Surgimiento de la Civilización en el Occidente de Yucatán: los Orígenes de la Complejidad Social en Sihó. Informe de Actividades de la Temporada de Campo 2001. Universidad Autónoma de Yucatán, Facultad de Ciencias Antropológicas, Mérida, Yucatán.
EA.FLMNH
2003 Colección fotográfica de 243 especies endémicas de la región maya, efectuada por Chr. Götz y en posesión del departamento de Arqueología Medio Ambiental del Florida Museum of Natural History, Vol. 2.
Emery, Kitty F.
2003 The Noble Beast: Status and Differential Access to Animals in the Maya World. World Archaeology 34(3):498-515.
Fisher, John W.
1995 Bone Surface Modifications in Zooarchaeology. Journal of Archaeological Method and Theory 2(1):7-68.
Folan, William, Abel Morales, José A. Hernández, Raymundo González, Lynda Florey Folan, David Bolles, Joel D. Gunn y Rosario Domínguez
2004 Recientes excavaciones en el antiguo barrio de Pozo del Monte, Las Mercedes en la ciudad y puerto de Champoton (Chakan Putun) Campeche: un lugar central del preclásico medio a posclásico en la costa oeste de la península de Yucatán y su corredor ecoarqueológico e histórico. En Los Investigadores de la Cultura Maya, 2005, pp.38-53. Universidad Autónoma de Campeche, México.
Forsyth, Donald y Aaron Jordan
2005 Reflexiones sobre la ocupación postclásica en Champoton a través de la cerámica. En Los Investigadores de la Cultura Maya, 2005, pp.32-37.Universidad Autónoma de Campeche, México.
De la Garza, Mercedes
1983 Relaciones Histórico-Geográficos de la Gobernación de Yucatán (Mérida, Valladolid y Tabasco). Instituto de Investigaciones Filológicas, Universidad Nacional Autónoma de México.
De Landa, Fray Diego
2003 Relación de las cosas de Yucatán. Ediciones Cien de México, México.
Götz, Christopher M
2007 Die Verwendung von Wirbeltieren durch die Mayas des nördlichen Tieflandes während der Klassik und Postklassik (600–1500 n.Chr.). Editorial Marie Leidorf, Serie Internationale Archäologie.
Hamblin, Nancy L.
1984 Animal Use of the Cozumel Maya. The University of Arizona Press, Tuscon, Arizona.
Howell, Steve N.G. y Sophie Webb
2004 A Guide to the Birds of Mexico and Northern Central America. Oxford University Press, New York.
Lee, Julian C.
2000 A Field Guide to the Amphibians and Reptiles of the Maya World. Cornell University Press, Ithaca and London.
Lee, Thomas A.
1985 Los Códices Mayas. Universidad Autónoma de Chiapas.
Leopold, A. Starke
2000 Fauna Silvestre de México. Editorial Pax, México.
Lyman, R. Lee
2000 Vertebrate Taphonomy. Cambridge Manuals in Archaeology, Cambridge University Press.
Márquez Morfín, Lourdes
1991 La dieta maya prehispánica en la costa yucateca. Estudios de la Cultura Maya 18:359-394.
O’Connor, Terry
2000 The Archaeology of Animal Bones. Texas A&M University Press, College Station.
2003 The Analysis of Urban Bone Assemblages: A Handbook for Archaeologists. The Archaeology of York, Principles and Methods 19/2, Counsil for British Archaeology.
Pollock, H.D.E. y Clayton E. Ray
1957 Notes on Vertebrate Animal Remains from Mayapan. Current Reports, Carnegie Institution of Washington, Department of Archaeology 41:633-656.
Pumajero, P.G. y F. Bernaldo de Quirós
1990 Huellas Humanas en Huesos – Análisis de sus implicaciones económicas. Revista de Arqueología 11(109):14-24. Madrid.
Reitz, Elizabeth y Elizabeth Wing
1999 Zooarchaeology. Cambridge Manuals in Archaeology, Cambridge University Press.
Reixach, Joan
1986 Huellas antrópicas. Metodología, diferenciación y problemática. Revista de Arqueología 7(60):6-14. Madrid.
Robins, Richard, Charleton Ray y John Douglas
1986 A Field Guide to Atlantic Coast Fishes. North America. Houghton Miffin Company.
Sierra Sosa, Thelma N.
1999 Xcambo. Codiciado enclave económico del Clásico Maya. Arqueología Mexicana 7(37):40-47. México.
Sierra Sosa, Thelma y Ángel Martínez
2001 Los entierros de Xcambo y sus implicaciones sociales. I’Inaj. Semilla de Maíz No.12:5-12. México.
Schmidt, Peter J.
2003 Informe del Proyecto Arqueológico Chichen Itza de julio de 1999 a diciembre de 2002. CRY INAH. (Informe no publicado).
2005 Nuevos hallazgos en Chichen Itza. Arqueología Mexicana 13(76):48-57. México.
Steadman, David W
1980 A Review of the Osteology and Paleonthology of Turkeys (Aves: Meleagridinae). Contributions in Science, Natural History Museum of Los Angeles County No.330:131-207. Los Ángeles.
Valadez Azúa, Raúl
2003 La domesticación animal. Instituto de Investigaciones Antropológicas, Universidad Nacional Autónoma de México.
White, Tim
1992 Prehistoric Cannibalism at Mancos 5MTUMR- 2346. Princeton University Press. USA.
Wing, Elizabeth S. y David Steadman
1980 Vertebrate Faunal Remains from Dzibilchaltun. En Excavations at Dzibilchaltun, Yucatan, Mexico (editado por E. W. Andrews IV y E.W. Andrews V), pp.331. Middle American Research Institute, Tulane University, New Orleans, Publicación No.48.
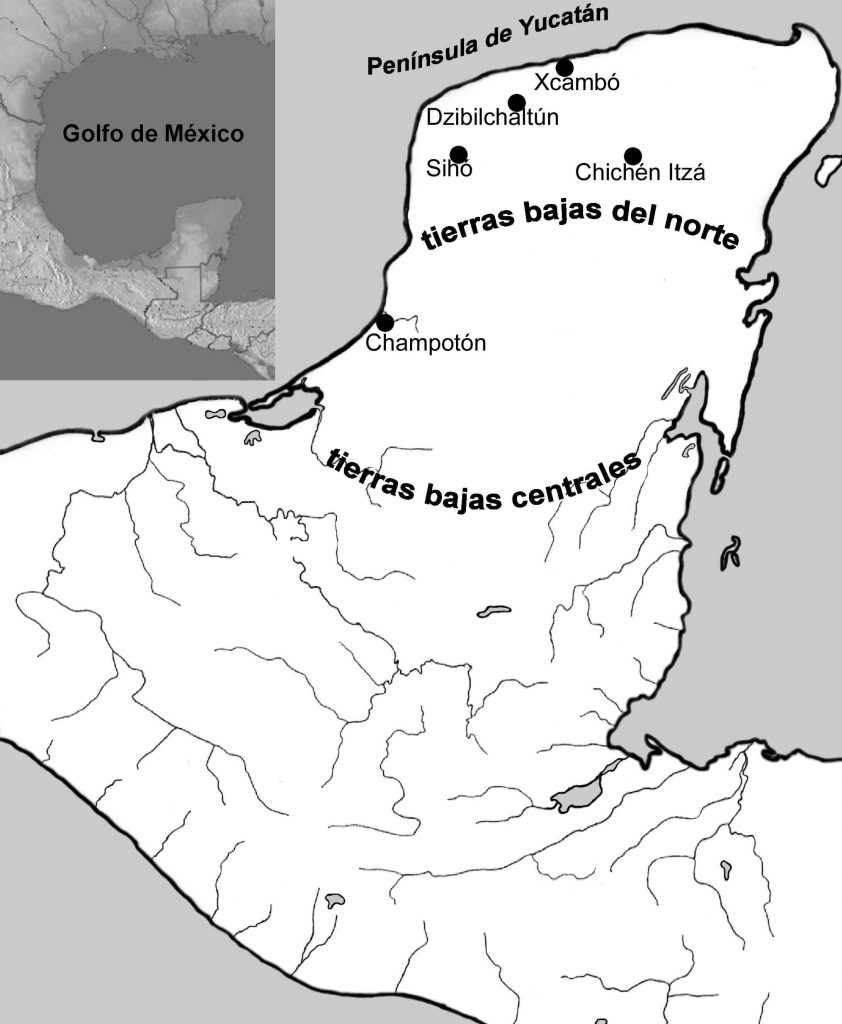
Figura 1 Mapa de la península yucateca, indicando los sitios investigados en este artículo

Figura 2 Tabla de presencia taxonómica (mamíferos y aves) de los cinco contextos y sitios comparados

Figura 3 Tabla de presencia taxonómica (reptiles y peces) de los cinco contextos y sitios comparados

Figura 4 Gráfica presentando las frecuencias taxonómicas del sitio de Xcambo, resumidas por tipos de animales. Las barras detallan el porcentaje ya sea del NISP o del MNI frente al total de la muestra

Figura 5 Gráfica presentando las frecuencias taxonómicas del sitio de Champoton, resumidas por tipos de animales. Las barras detallan el porcentaje ya sea del NISP o del MNI frente al total de la muestra

Figura 6 Gráfica presentando las frecuencias taxonómicas del sitio de Chichen Itza, resumidas por tipos de animales. Las barras detallan el porcentaje ya sea del NISP o del MNI frente al total de la muestra

Figura 7 Gráfica presentando las frecuencias taxonómicas del sitio de Dzibilchaltun, resumidas por tipos de animales. Las barras detallan el porcentaje ya sea del NISP o del MNI frente al total de la muestra

Figura 8 Gráfica presentando las frecuencias taxonómicas del sitio de Siho, resumidas por tipos de animales. Las barras detallan el porcentaje ya sea del NISP o del MNI frente al total de la muestra

Figura 9

Figura 10